







研究成果のポイント
-
均一な形状やサイズを持つ植物の萼片器官の形態が,相反する細胞成長の不均一性によって迅速に平均化され,形成される仕組みを,野生型シロイヌナズナによって解明。
-
器官の形やサイズが不均一になる遺伝子とその変異体を特定することに成功し,野生型と変異体の違いを調べることで,器官が成長を止める仕組みを発見。
-
先駆的なデータ解析手法によって,均一な器官形状を持つ野生型器官がより細胞成長に多様性を持つことを立証し,数理モデルによって均一な器官への細胞レベルの不均一性が果たす役割を解明。
-
自然界のあらゆる生物器官の形態形成メカニズムの解明が期待される。
研究成果の概要
北海道大学電子科学研究所(所長 西井準治教授)附属社会創造数学研究センターの津川暁特任助教,李振風准教授,小松崎民樹教授は,リヨン高等師範学校(フランス)のアレツキー・ボダード教授,コーネル大学(アメリカ)のエイドリアン・ロダー助教,マックス・プランク植物育種学研究所(ドイツ)のリチャード・スミス上級研究員らと共同で,発生生物学の長年の神秘である器官形状やサイズがほぼ同じになる性質(頑健性)※1について,シロイヌナズナの萼片器官の遺伝データや蛍光データを解析し,頑健性と相反する細胞成長のランダム性(不均一性)が形態形成にとって重要な役割を果たしていることを明らかにしました。これは自然界の植物器官(花びら,葉,根など)の正確な形状がよりミクロな細胞成長のランダム性を必要とするという,一見直感に反する巧妙な生命の仕組みが備わっている可能性を示唆しています。
この成果は,「ゾウはなぜ大きく,ネズミはなぜ小さいのか」「人間の左右の手はなぜほぼ同サイズなのか」などの発生生物学の創始以来の基本的課題に一石を投じるもので,植物器官に限らず広く自然界のあらゆる生物器官の形態形成メカニズムの解明に繋がるものと期待されています。
本研究成果は,国際的プロジェクト「ヒューマン・フロンティア・サイエンス・プログラム」(李准教授らによる受賞),及び北海道大学,東北大学,東京工業大学,大阪大学,九州大学の5附置研究所のネットワーク型による文部科学省「ナノマクロ物質・デバイス・システム創製アライアンス」「物質・デバイス領域共同研究拠点」などの支援を受けました。
論文発表の概要
研究成果の概要
(背景)
なぜゾウは大きく,ネズミは小さいのでしょうか。なぜ人間の2つの手はサイズが1%しか違わないのでしょうか。発生生物学は,一見単純そうなこれらの疑問に未だに答えることができていません。最も重要な未解決問題は,生命が正確なサイズや形状に到達するまでに,どのように成長を止めることを“知る”ことができるのか,という問題です。植物はほとんど同形の花を生成する性質があり,顕微鏡で観測することが比較的容易であるため,この問題に対して格好の研究題材といえます。これまでの先行研究では,すべての花を大きく(または小さく)するような遺伝子や,すべての細胞を大きく(または小さく)する遺伝子を特定することはできていましたが,植物がどのようにして成長を止めてほぼ同形のサイズや形状になっているかはよくわかっていませんでした。本研究の始まりは,人間の2つの手でいえば指紋や皮下脂肪がそれぞれ異なるように,植物器官の中の細胞形状やサイズが個々の花で異なるというミクロな観測結果が手掛かりとなりました。
(研究手法)
本研究では,生物学・遺伝工学・コンピューター科学・数理物理学などの広範な学問の連携によって,モデル植物であるシロイヌナズナの再生的な萼片器官の形成メカニズムの解明に挑戦しています。最も重要な手法は遺伝学的スクリーニング技術※2で,異なる表現型※3をもつ遺伝子を特定することができます。また,最先端の光学顕微鏡による細胞の分裂と成長の観測と,力学顕微鏡による器官内の物理的力を観測することで,萼片器官を生物・物理の両側面から詳細に測定しました。測定結果として得られた蛍光データに対して新しくイメージング技術や統計解析を開発して,細胞動態の定量的評価が可能になりました。
(研究成果)
顕微鏡による測定から,細胞成長の振る舞いが隣接細胞の成長と比べて著しく異なり,均一ではないことを発見しました。この発見をもとに,細胞成長方向が時間と共に変化するような器官成長の数理モデルを構築することを考えました。
数理モデルの着想を直感的に例えてみます。例えば,たくさんの人が空間に“でたらめに(不均一に)”置かれた目印に向かって長時間歩く場合は,すぐに皆が散らばってしまいますが,それぞれの人が数秒の間隔で別のでたらめな目印に向かって歩く向きを変えれば,人々は散らばらずに長時間ある場所にとどまっていることができます。
この着想を細胞の成長方向に応用し(人の歩く方向に対応),実際の萼片器官のデータで立証することに成功しました。モデルの整合性を立証するために,自然界に存在する野生型の萼片と遺伝学的スクリーニングによって特定された器官形状やサイズが不均一になる変異体の萼片を比較しました。驚くべきことに,バラバラな萼片サイズを持つ変異体の方が細胞成長の不均一性が小さいことが確認されました。活性酸素※4濃度を詳しく調べることで,器官サイズを決定する原因が遺伝子による活性酸素の制御であるということが判明しました。興味深いことに,抗酸化物として知られるビタミンCを投与すると花弁が大きくなることが遺伝子制御によって説明することができます。
(今後への期待)
本研究は,器官発生が細胞動態の巧みな組織化の連鎖によって起こると信じられてきた従来からの発生生物学の基本問題に挑戦し,生命体の正常な機能の一つとして無秩序から秩序を創出する仕組みを提案しています。これまで観測の邪魔をすると考えられてきた細胞レベルでのノイズやゆらぎを,今後はむしろ積極的に調べていくことで生命の形態形成に潜むメカニズムを明らかにしていくことができると考えています。また,本研究の考え方は植物器官以外の生命体にも適用することができるため,研究が進んでいるショウジョウバエの羽や大腸菌の形態形成への応用に加え,人間を含むより高度な多細胞生物のサイズや形状を決める基本原理となることが期待されています。
お問い合わせ先
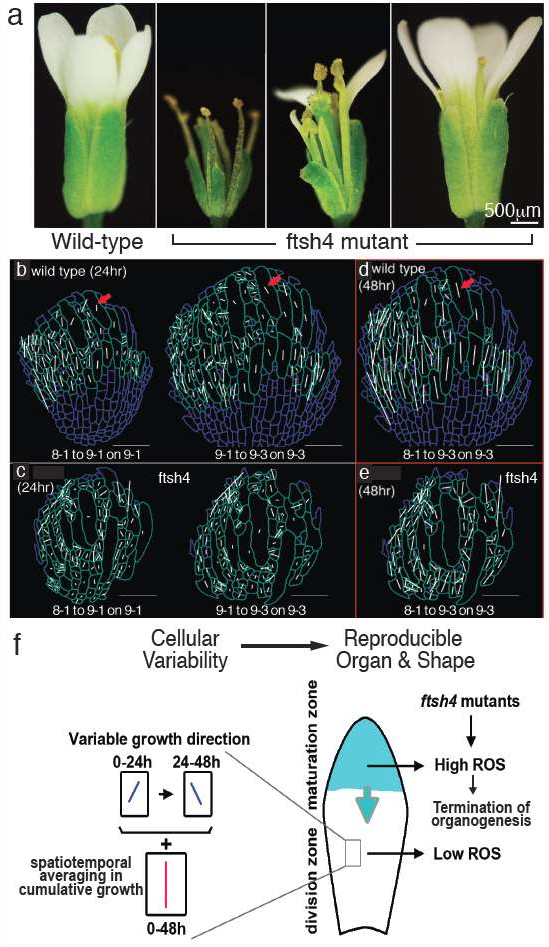
図 1 (a)成熟後の野生型(左側)及びftsh4 変異体 ※5 (右)。 ftsh4 変異体の花は同じ花序のものだが,その萼片のサイズは多様である。
(b)-(c)主成長方向 ※6 における空間的変動。野生型の主成長方向(PDG;白線)(b)と 24 時間間隔での成長を計算したftsh4 (c)の萼片細胞。萼片細胞が示す 24 時間の成長間隔で の最大成長方向における時空間変動(矢印)。白線の方向は最大成長の方向,白線の長さはその方向に おける成長の規模を示している。解析では成長線(白線)が表示された細胞のみを用いた。
(d)-(e)48時間にわたる時間平均化。0–48時間の累積成長のPDGを計算した。野生型の萼片細胞の48時間の最大成長方向(d)は24時間の最大成長方向(b)における積算結果であり,より協調的である。ftsh4の萼片細胞(e)は,48時間で統合されたPDGが平行になっていない中で,野生型よりも時間平均化の変動性が少ない。
(f)細胞成長は変動性が高く,時間によって変化する。例えば,細胞の24時間ごとのPDGは左に傾いた後,右に傾くかもしれない。時間及び空間による変化は細胞変動を平均的にするため,例えば 48時間による累積成長では均一な器官を非常に規則的に作り出す。活性酸素(右図の萼片上部)が細胞の成熟,細胞分裂の縮小,及び成長の停止を促進する。成熟した細胞において,ROSの蓄積は先端で始まり底に向かって降下していく(右図の萼片内部の矢印)。ftsh4変異体が原因で,より遅くかつ変動性の少ない細胞成長,細胞分裂の縮小,成長の時空間平均化の抑制,及び萼片サイズにおける頑健性の喪失をもたらすROSの増加,変動,早期の蓄積が起こっている。
- 1.頑健性:
-
様々な擾乱に抗って機能を強靭かつ健全に維持する特性のこと。環境の変化に対して,ダメージや変性,機能損失を最小限にとどめながら処理することが出来るとき,そのシステムや生物は「頑健である」と言える。
- 2.遺伝学的スクリーニング:
-
ある表現型に関わる遺伝子を順遺伝学的方法と逆遺伝学的方法により発見すること。順遺伝学的方法とは遺伝性がみられる形質(表現型)からその原因となる遺伝子を探り当てる研究を指し,逆遺伝学的手法とは着目した遺伝子の発現を抑制することによって起こる表現型の変化を調べ,その遺伝子の機能を解析する研究を指す。本研究では両方法を組み合わせて変異型の特定を行っている。
- 3.表現型:
-
生物の発生,形態形成,生理学的性質,物理化学的振る舞いなど一連の過程を経て観測される特徴や形質のこと。つまり,その形質において観察される個々のタイプ(型)を指す言葉である。例えば,「インゲン豆の形」という形質の中に,「丸い」「白い」という表現型があり,「果実が甘い」「病気に対する抵抗性」など特定の条件や分析の結果で判明するような形質も,遺伝子型が形質として実現された表現型という。
- 4.活性酸素
-
活性酸素は,酸素分子(O2)よりも活性化された状態の酸素分子とその関連物質を指す。活性酸素分子種としてはスーパーオキシド(O2–),ヒドロキシルラジカル(HO),過酸化水素(H2O2),一酸化窒素(NO)などがある。活性酸素は,血管を障害し,老化や癌化を促進する作用を持つことが知られており,脂質,特に,細胞膜のリン脂質を酸化させたり,たんぱく質やDNAに酸化障害を与える。
- 5.ftsh4 変異体:
-
AAA-ATPaseとメタロプロテアーゼ(活性中心に金属イオンが配座しているたんぱく質分解酵素)をもつ膜たんぱく質プロテアーゼの一つであるFtsH4というたんぱく質に変異導入したftsh4変異体を特定した。ftsh4変異体は活性酸素を増加し,葉の形態異常を引き起こすことがすでに知られている。
- 北海道大学: http://www.hokudai.ac.jp/news/160712_es_pr.pdf
- コーネル大学: http://www.news.cornell.edu/stories/2016/07/flower-bud-uniformity-beholden-time-and-space
- リヨン師範学校
- フランス語: http://www.ens-lyon.fr/zoom-sur-/variable-cell-growth-yields-reproducible-organ-development-through-spatiotemporal-averaging-316532.kjsp?RH=ENS-LYON-FR-RECH-FAI
- 英語: http://www.ens-lyon.fr/en/variable-cell-growth-yields-reproducible-organ-development-through-spatiotemporal-averaging-316832.kjsp?RH=TEMP-INTER-EN
